ГЕТЕРОЗИС ГЕТЕРОЗИС
(от греч. heteroiosis - изменение, превращение), «гибридная мощность», превосходство гибридов по ряду признаков и свойств над родительскими формами. Термин «Г.» предложен Дж. Шеллом в 1914. Как правило, Г. характерен для гибридов первого поколения, полученных при скрещивании неродственных форм: разл. линий, пород (сортов) и даже видов. В дальнейших поколениях (скрещивание гибридов между собой) его эффект ослабляется и исчезает. Гипотеза «сверхдоминирования», или моногенного Г., предполагает, что гетерозиготы по определ. гену превосходят по своим характеристикам соответствующие гомозиготы. К явлению, иллюстрирующему эту гипотезу, можно отнести межаллельную комплементацию. В основе ряла др. гипотез лежит предположение о наличии у гибрида большего числа доминантных аллелей разных генов по сравнению с родительскими формами и о взаимодействии между этими аллелями. Гипотезы синтетического характера основаны как на внутригенных, так и на межгенных взаимодействиях. О значении гетерозиготности как основы Г. свидетельствует и тот факт, что в природных популяциях особи гетерозиготны по большому числу генов. Более того, в гетерозиготном состоянии сохраняется мн. аллелей, проявляющих в гомозиготном состоянии неблагоприятные эффекты на жизненно важные признаки. Г. имеет важное значение в с.-х. практике (у с.-х. животных и растений Г. нередко приводит к значит, повышению продуктивности и урожайности: получение простых и двойных межлинейных гибридов кукурузы позволило повысить на 20-30% валовые сборы зерна), однако его использование часто недостаточно эффективно, т. к. до сих пор не решена проблема закрепления Г. в ряду поколений. В качестве подходов к решению этой проблемы рассматривается вегетативное размножение гетерозисных форм, полиплоидия и разл. нерегулярные формы полового размножения (апомиксис, партеногенез и др.).
.(Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. - 2-е изд., исправл. - М.: Сов. Энциклопедия, 1986.)
гетеро́зис (гибридная мощность, гибридная сила), превосходство гибридов первого поколения над родительскими формами по жизнеспособности, урожайности, плодовитости и ряду других признаков. Для получения эффекта гибридной мощности важно в качестве родителей выбирать неродственные формы, представляющие различные линии, породы, даже виды. На практике наилучшие родительские пары, дающие наиболее ценные гибриды, отбираются в результате многочисленных скрещиваний, позволяющих выявить наиболее удачную сочетаемость различных линий. При скрещивании между собой следующих поколений гетерозис ослабевает и затухает.
В основе гетерозиса лежит резкое повышение гетерозиготности у гибридов первого поколения и превосходство гетерозигот
по определённым генам над соответствующими гомозиготами
. Таким образом, явление гибридной мощности противоположно результату близкородственного скрещивания – инбридинга
, имеющему для потомства неблагоприятные последствия. Генетический механизм гетерозиса (он до конца не выяснен) связывают также с наличием у гибрида по сравнению с родителями большего числа доминантных генов, взаимодействующих между собой в благоприятном направлении.
Гетерозис широко используется в практике сельского хозяйства для повышения урожайности с.-х. культур и продуктивности с.-х. животных. В 1930-е гг. селекционеры США резко повысили урожайность кукурузы, применяя гибридные семена. Одна из важных задач селекции
– поиски путей «закрепления» гетерозиса, т.е. сохранения его в ряду поколений.
.(Источник: «Биология. Современная иллюстрированная энциклопедия.» Гл. ред. А. П. Горкин; М.: Росмэн, 2006.)
Синонимы :
Смотреть что такое "ГЕТЕРОЗИС" в других словарях:
Ускорение роста, увеличение размеров, повышение жизнестойкости и плодовитости гибридов первого поколения по сравнению с родительскими формами растений или животных. Обычно во втором и последующих поколениях гетерозис затухает. Гетерозис широко… … Финансовый словарь
Гетерозис (в переводе с греческого языка изменение, превращение) увеличение жизнеспособности гибридов вследствие унаследования определённого набора аллелей различных генов от своих разнородных родителей. Это явление противоположно инбредной … Википедия
- (от греческого heteroiosis изменение, превращение), свойство гибридов первого поколения превосходить по жизнестойкости, плодовитости и другим признакам лучшую из родительских форм. Во втором и последующих поколениях гетерозис обычно затухает.… … Современная энциклопедия
В селекции животных и растений особое место занимает явление гибридной мощности, или гетерозиса , которое заключается в следующем.
При скрещивании разных рас, пород животных и сортов растений, а также инбредных линий гибриды F 1 по ряду признаков и свойств часто превосходят исходные родительские организмы. Скрещивание гибридов между собой ведет к затуханию этого эффекта в следующих поколениях. В настоящее время гетерозис установлен для всех изученных видов.
Хотя эффект гетерозиса известен с древнейших времен, его природа до сих пор остается мало изученной. Первое толкование биологического значения гетерозиса в эволюции животных и растений и попытка объяснить механизм этого явления принадлежали Ч. Дарвину. Он систематизировал большое количество фактов и сам ставил опыты по изучению гетерозиса. По мнению Ч. Дарвина, гетерозис служит одной из причин биологической полезности скрещивания в эволюции видов. Перекрестное оплодотворение поддерживается естественным отбором именно потому, что оно служит механизмом сохранения наибольшей гетерозисности,
Глубокий научный анализ явления гетерозиса стал возможен только с начала XX в. после открытия основных генетических закономерностей. При рассмотрении гетерозиса мы будем касаться возникновения его при скрещивании преимущественно инбредированных линий одного вида, поскольку в этом случае легче выяснить его генетический механизм.
С начала нашего столетия на кукурузе стали проводить систематическое исследование скрещиваний между инбредными линиями. При этом Г. Шеллом было показано, что скрещивание некоторых линий дает гибридные растения, более урожайные по зерну и вегетативной массе, чем исходные линии и сорта. Приведены опытные данные, показывающие низкую урожайность инбредных линий, значительное повышение урожайности в F 1 и снижение в F 2 при самоопылении растений F 1 .
За последние 30 лет в ряде стран, в том числе и в нашей, посев гибридными семенами стал основным приемом производства кукурузы как на зерно, так и для силоса. Для получения гибридных семян сначала создают инбредные линии из лучших сортов, отвечающих требованиям данного климатического района (инбредная линия создается в течение 5-6 лет путем самоопыления). При отборе линий оценивают их качества и свойства, которые необходимо получить у будущего гибридного организма. Инбридинг в линиях не может быть эффективным, если он не сопровождается отбором.
Создав большое число инбредных линий, приступают к скрещиванию между ними. Межлинейные гибриды первого поколения оценивают по эффекту гетерозиса ; исходя из этого показателя отбирают линии с лучшей комбинационной ценностью и затем размножают их в больших масштабах для производства гибридных семян. Работа по созданию инбредных линий и оценке их комбинационной ценности постоянно ведется в селекционных учреждениях. Чем больше создано ценных линий, тем вернее й скорее можно отыскать лучшие гибридные комбинации с необходимым сочетанием свойств.
При получении гибридных семян для производственных целей исходные линии, дающие при скрещивании наибольший эффект гетерозиса, высевают рядами, чередуя материнские и отцовские формы. Чтобы обеспечить опыление между ними, с материнских растений удаляют мужские соцветия (метелки). Теперь разработана новая схема производства гибридных семян с использованием цитоплазматической мужской стерильности, что позволило значительно сократить труд на удаление метелок у растений материнской линии. Таким путем получают простые межлинейные гибриды кукурузы. Этот метод в принципе является общим для семеноводства гибридов различных перекрестноопыляющихся растений.
В настоящее время в практике сельского хозяйства простые межлинейные гибриды кукурузы не используются, так как затраты на получение таких семян не окупаются. Теперь широко внедряется в практику посев семян двойных межлинейных гибридов. Последние получают путем скрещивания двух простых гибридов, проявляющих гетерозис.
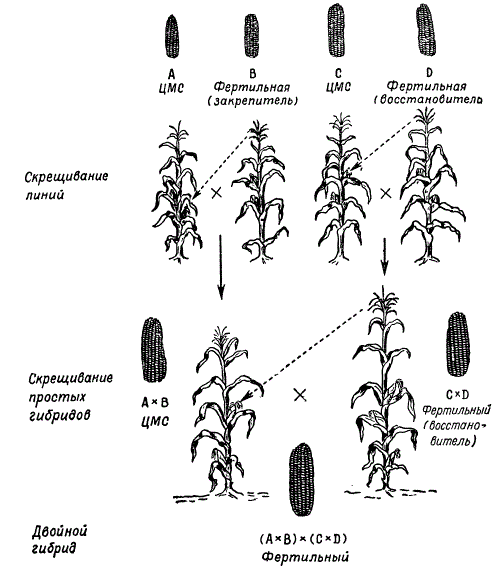
По данным М. И. Хаджинова и Г. С. Галеева, результаты оценки продуктивности разных гибридов - межсортовых, сортолинейных и двойных межлинейных - показывают, что наиболее продуктивны двойные межлинейные гибриды.

Подбор простых гибридов для получения наиболее продуктивных двойных гибридов является важным этапом их селекции. Лучшие результаты дает скрещивание линий, происходящих из различных сортов. Так, например, если один простой гибрид получен от скрещивания инбредных линий двух сортов А X В, а другой - от скрещивания линий других сортов С х D, то двойной гибрид (А X В) х (С X D) дает гетерозис чаще, чем если бы двойной гибрид был получен от скрещивания простых гибридов, происходящих от линий одного сорта: (А х А 1 х (А 2 х А 3) или (В х B 1) х (В 2 х В 3).
Для успешного развития гибридного семеноводства необходимо исследовать, во-первых, как долго следует вести инбридинг, чтобы получить гомозиготные линии, и, во-вторых, разработать методы более быстрой оценки их комбинационной ценности.
Все, что было сказано в отношении кукурузы, по-видимому, является общим для получения гибридов у других высших перекрестноопыляющихся растений, а также у животных. В настоящее время в птицеводстве и в свиноводстве ряда стран широко используется скрещивание инбредных линий, происходящих из одной или разных пород. Следует особо подчеркнуть, что широкое использование гибридов в животноводстве возможно только при высоком уровне племенной работы и наличии ценных пород, то совершенно ясно, что в большинстве случаев инбредные линии в среднем будут иметь всегда более низкие показатели, чем сорта. О наличии гетерозиса следует говорить лишь в том случае, когда межлинейный гибрид превосходит не только родителей (линии), но и сорта, или породы, от которых произошли эти линии.
Как мы знаем, гены детерминируют свойства организма на всех стадиях онтогенеза начиная с момента оплодотворения. Гены ядра ооцита способны детерминировать свойства цитоплазмы яйцеклетки еще до оплодотворения. Характер реализации генотипа изменяется в зависимости от свойств цитоплазмы зиготы. Таким образом, проявление гетерозиса у гибрида зависит также и от свойств цитоплазмы. Роль цитоплазмы в определении гетерозиса доказывается следующим образом. При реципрокном скрещивании двух линий А X В и В х А гетерозис по одним и тем же свойствам нередко проявляется у гибридов только одного из скрещиваний и не проявляется у гибридов другого.
Проявление гетерозиса обусловлено индивидуальным развитием гибрида. В онтогенезе он реализуется неравномерно. На одних стадиях онтогенеза проявляется гетерозис по одним признакам, а на других - по другим или только по некоторым. Так, в раннем возрасте у одного и того же гибрида может наблюдаться гетерозис в отношении скорости роста отдельных частей организма и повышенной устойчивости к заболеваниям, но его может не быть, например, в отношении устойчивости к неблагоприятной температуре. Гетерозис по этому свойству может проявляться позднее.
Сильное влияние на проявление гетерозиса оказывают также факторы среды, в которых развивается гибридный организм. Есть основание считать, что гетерозис по жизнеспособности и устойчивости к заболеваниям лучше выявляется при неблагоприятных для развития условиях среды. Еще Ч. Дарвином было высказано предположение, что гетерозис у гибридов обусловлен более широкой нормой приспособительных реакций. Исследования последних лет показали, что гибриды F 1 в сравнении с инбредными линиями характеризуются большим разнообразием процессов обмена, участием большего числа различных метаболитов, ростовых веществ и энзимов. Гетерозис проявляется не только в целом организме, но и на клеточном уровне.
Выяснение генетических механизмов гетерозиса остается нерешенной проблемой. В настоящее время имеются три гипотезы, пытающиеся объяснить возникновение гетерозиса:
- гетерозиготное состояние по многим генам,
- взаимодействие доминантных благоприятных генов,
- сверхдоминирование - гетерозигота превосходит гомозиготы.
Как мы уже говорили, при скрещивании гомозиготных инбридированных линий гибриды первого поколения по многим генам приобретают гетерозиготное состояние. При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей. Схематически это можно представить так: одна инбредная линия в гомозиготном состоянии имеет рецессивную аллель одного гена ааВВ, а вторая - другого гена ААbb. Каждая из этих рецессивных аллелей генов в гомозиготном состоянии определяет какую-нибудь недостаточность, которая снижает жизнеспособность инбредной линии. При скрещивании линий ааВВ X ААbb у гибрида объединяются доминантные аллели обоих генов (АаВb). Гибриды F 1 проявят при этом по указанным генам не только гетерозис, но и единообразие. В F 2 число особей с двумя доминантными генами в гетерозиготном состоянии будет лишь 4/16, поэтому гетерозисными оказываются не все особи. В дальнейших поколениях число гетерозигот сокращается, а число гомозигот увеличивается. В силу этих причин гетерозис в последующих поколениях затухает. Такова схема одной из генетических гипотез гетерозиса.
Мы привели пример с двумя генами, но физиологические свойства определяются огромным числом генов. Кроме того, неверно считать, что доминантные аллели всегда положительно, а рецессивные отрицательно влияют на жизнеспособность организма и его продуктивность. Доминантные аллели дикого типа чаще оказывают благоприятное действие, чем рецессивные. Это можно объяснить тем, что неблагоприятные доминантные мутации подвергаются более жесткому отбору - они уже в зиготе и на эмбриональных стадиях подвергаются элиминации, и отбором сохраняются лишь благоприятные. И так как само свойство доминирования гена эволюционирует под контролем отбора, то аллели дикого типа и оказываются более благоприятными для организма. Поэтому подбор в гибридной комбинации доминантных аллелей может скорее обеспечить гетерозис. Если бы гетерозис обусловливался простым набором доминантных аллелей, имеющихся в популяции, то этот набор было бы легко составить путем ряда скрещиваний и получить гетерозисные сочетания. Весьма возможно, что часть пород и сортов выводилась именно путем скрещивания и комбинаций линий, имеющих набор благоприятных доминантных аллелей. Но до сих пор не удавалось закрепить гетерозис в гибридной комбинации F 1 , т. е. получить не расщепляющиеся в F 2 формы.
Д. Джонсом еще в 1917 г. было предложено дополнение к изложенной гипотезе. Согласно Д. Джонсу, разные гены, дающие в сочетании гетерозис, находятся в пределах одной группы сцеплений: например, AbcdE - в одной, а соответственно aBCDe - другой гомологичной хромосоме. Именно это обстоятельство, с точки зрения Д. Джонса, затрудняет отбор в F 2 полностью гомозиготных форм по благоприятным доминантным генам, дающим сочетании гетерозис. Для осуществления такого сочетания доминантных аллелей необходимо, чтобы в этой паре хромосом AbcdE//aBCDe произошел минимум двойной кроссинговер, который привел бы к возникновению хромосомы с одними доминантными аллелями ABCDE. Малая вероятность одновременного осуществления такого события в нескольких хромосомах и является, по мнению Д. Джонса, причиной, затрудняющей закрепление гетерозиса. Гибрид же F 1 содержит все эти доминантные аллели, и поэтому у него наблюдается гетерозис.
Такова схема объяснения гетерозиса согласно второй гипотезе - гипотезе взаимодействия доминантных благоприятных генов, которую называют иногда также гипотезой набора кумулятивных доминантных аллелей, а сам гетерозис при этом - мутационным. Эта гипотеза в целом исходит из представления о простом суммировании эффекта доминантных аллелей с комплементарным действием.
Третья гипотеза исходит из того, что гетерозиготное состояние аллелей имеет превосходство над гомозиготным (АА < Aa > аа). Здесь можно допустить благоприятное трансположение аллелей дикого типа и мутантных аллелей, которое каким-то образом усиливает эффект действия генов. Указанное объяснение гетерозиса называют гипотезой сверхдоминирования.
Ни одна из трех гипотез не может считаться единственно правильной. Сейчас преждевременно отдавать предпочтение той или другой. Вероятно, они все окажутся правильными, но для разных случаев. Наверное, каждый из механизмов, предусматриваемых этими гипотезами, играет роль в определении гибридной мощности. Гетерозис является сложным явлением как по механизму возникновения, так и по проявлению его в онтогенезе.
Очевидно, окончательный вывод о генетических механизмах гетерозиса можно будет сделать лишь после того, как раскроется картина взаимодействия генов в генетической системе на биохимическом и молекулярном уровнях. Как мы уже знаем, подавление проявления мутаций с помощью супрессоров позволяет исправлять нарушения в действии гена и его аллелей. Возможно, что явление гетерозиса есть суммарный результат работы не основных генов, определяющих развитие признаков организма, а набора супрессирующих генов в генотипе. Важным моментом изучения гетерозиса становится исследование плазменных отношений.
Основной задачей использования гетерозиса в селекции является закрепление его, т. е. сохранение эффекта гетерозиса в процессе воспроизведения гибрида. Решение этой задачи мыслится в нескольких аспектах: во-первых, закрепление гетерозиса путем перевода гибридного организма с полового размножения на апомиктическое, что, по-видимому, возможно для некоторых растений; во-вторых, закрепление гетерозиса путем перевода диплоидного гибрида, проявляющего гетерозис, в полиплоидное состояние. В этом случае гетерозиготная комбинация генов будет сохраняться дольше.
У вегетативно размножающихся растений поддержание ценных гибридных комбинаций, полученных половым путем, осуществляется вегетативным размножением (черенками, прививками, клубнями и т. д.).
Существует ряд других путей поддержания гетерозиса в ряду поколений, однако все они еще недостаточно разработаны и проверена.
Способ предназначен для использования в сельском хозяйстве. Цель - значительное повышение эффективности гибридизации сельскохозяйственных культур. Гетерозис гибридов проявляется только в первом поколении. Установлено, что затухание гетерозиса в последующих поколениях гибрида в основном происходит из-за перехода рецессивных леталей, полулеталей и субвиталей в гомозиготное состояние и нарушения комплекса благоприятных скоординировано действующих генов. Устранение этих явлений приводит к закреплению гетерозиса в последующих поколениях. Оно выполняется посредством возвратных скрещиваний гибрида с искусственно полученными от него абсолютно гомозиготными андрогенетическими сыновьями, после чего генетически трансформируемый гибрид практически полностью очищается от вредных генов и одновременно сохраняет в целости комплекс благоприятных генов, определяющих гетерозис. Это позволяет полностью сохранить гетерозис в последующих уже промышленных поколениях, полученных в результате несложных внутригибридных скрещиваний, что доказано экспериментами на тутовом шелкопряде. Способ также предназначен для сельскохозяйственных растений, у которых возможно получение андрогенетических абсолютно гомозиготных особей. 3 з.п.ф-лы, 4 ил.
Изобретение относится к способам, используемым в сельском хозяйстве. Общеизвестен естественный способ сохранения гетерозиса в ряду последующих поколений посредством вегетативного размножения у растений, дополнительно обладающих половым размножением. Многочисленные исследования в этой области на других растениях, не способных к вегетативному размножению, и животных не увенчались полным успехом (1), потому что природа гетерозиса все еще оставалась большой загадкой генетики (2). В литературе не было высказано даже сколько-нибудь реальных теоретических подходов к кардинальному решению этой важной проблемы. У некоторых животных гетерозис можно закрепить клонированием. Однако этим способом пока получают единицы идентичных матери потомков. У тутового шелкопряда клонирование разработано более успешно, но для практического использования в плане сохранения гетерозиса оно не приемлемо по двум причинам: из-за большой трудоемкости массового получения партеногенетического потомства и меньшей, по сравнению с самцами, продуктивности женского пола, из которого состоят клоны (3). Перспективные результаты получены авторами после того, как они разработали метод мейотического партеногенеза и получили от партеногенетических клонов абсолютно гомозиготных самцов тутового шелкопряда (4). Их возвратное скрещивание с партеногенетическим клоном гибридного происхождения позволило закреплять гетерозис в беккроссных поколениях (5). Но это было открытием лишь принципиальной возможности закрепления гетерозиса. Практического же значения этот метод не имел и, следовательно, не мог быть запатентован в качестве способа. Это объяснялось тем, что гомозиготных самцов могли получать только от высокожизнеспособных женских партеноклонов с высокой склонностью к партеногенезу. У коммерческих пород и гибридов абсолютных гомозиготов практически получать не удавалось, поэтому мейотический партеногенез был использован только для рекогносцировочных опытов, направленных на выяснение возможности решения проблемы. Изобретение способа закрепления гетерозиса у тутового шелкопряда, пригодного для производства, стало возможным после открытия авторами односпермичного андрогенеза (1998, неопубликовано). Сущность изобретения. Гетерозис проявляется только в первом поколении гибрида. В последующих поколениях, начиная со второго, он резко затухает. Поэтому, чтобы вырастить гетерозисный гибрид приходится каждый раз вновь повторять межсортовую или межпородную гибридизацию. Этот процесс технически сложный и весьма трудоемкий, а применительно ко многим растительным культурам он просто невыполним, хотя их гибриды, если бы они были получены, дали бы удивительно высокие урожаи по сравнению с родительскими формами. Примером этого служат многие сельскохозяйственные растения. Эти проблемы были бы кардинально решены, если бы удалось разработать эффективный способ закрепления гетерозиса в последующих поколениях. Такой способ одновременно открыл бы совершенно новый подход к созданию еще более выдающихся по гетерозису гибридов. Известно, что любой промышленный гибрид получают посредством скрещивания огромной массы индивидуумов двух родительских форм. А эти индивидуумы весьма дифференцированы по комбинационной способности. Поэтому производство довольствуется средним гетерозисом по всем вместе взятым индивидуальным гибридам, каждый из которых происходит от половых клеток двух родителей. В то время как редкие индивидуальные гибриды обладают поистине фантастическим гетерозисом, в следующем поколении он безвозвратно теряется. Предлагаемый способ позволит закреплять этот мощный гетерозис в последующих поколениях гибрида и размножать его в неограниченных количествах. Одной из причин гетерозиса считали благоприятное действие на развитие и жизнедеятельность организма гетерозиготности вообще всех генов, независимо от их специфичности (гипотеза "сверхдоминирования"). Авторы на тутовом шелкопряде экспериментально доказали, что гетерозис возникает в результате двух главных причин. Первая - интеграция в генотипе гибридов большого количества скоординированных в своем действии благоприятных генов, контролирующих жизнеспособность. Вторая - переход в гетерозиготное состояние не всех генов генотипа, а только рецессивных деталей, полулеталей и субвиталей (4). На фиг. 1 приведены доказательства этого. Следовательно, снижение гетерозиса в последующих поколениях гибридов в основном объясняется неизбежным при скрещивании гибрида в его пределах переходом части рецессивных деталей и полулеталей в гомозиготное состояние и нарушением в процессе мейоза комплекса благоприятных генов, повышающих жизнеспособность. Поэтому авторы пришли к выводу, что закрепить гетерозис в последующих поколениях можно, если в генотипе гибрида полностью сохранить или даже улучшить комплекс всех благоприятных генов и практически полностью удалить из генотипа рецессивные летали и полулетали. Эта задача решена авторами следующим способом. В качестве исходного материала выбирают генетически отдаленные две породы, от скрещивания которых возникают наиболее высокогетерозисные гибриды. От этих двух пород получают серию индивидуальных гибридов, каждый из которых происходит только от двух родителей. Путем сравнительных испытаний выбирают 10 лучших по гетерозису индивидуальных гибридов. От каждого гибрида получают абсолютно гомозиготных потомков методом односпермического андрогенеза, выполнение которого доступно селекционерам. Для этого неосемененных самок любой породы облучают - лучами в дозе 80 кр. Затем самки спариваются с самцами индивидуальных гибридов. Отложенные яйца в возрасте 60-80 минут после откладки при температуре 25 o C прогревают 210 минут в воде, нагретой до 38 o C. Абсолютные гомозиготы в подавляющем большинстве погибают на разных стадиях развития из-за того, что в гаплоидном генотипе, доставшимся им от отца, содержится много летальных, полулетальных и субвитальных генов. При диплоидизации ядра пронуклеуса они переходят в гомозиготное состояние, чаще всего несовместимое с нормальным развитием организма. Выживают только те гомозиготы, которым в ходе мейоза не досталось или досталось, но очень мало, вредных генов, в основном слабого действия (5). Выращенных абсолютно гомозиготных особей возвратно (беккросс) скрещивают с исходным гибридом, получая, таким образом, первое беккроссное поколение (фиг. 2). Созревание исходного гибрида и абсолютных гомозиготов должно быть синхронизировано путем задержки начала выращивания первого на время, равное продолжительности цикла развития взятого объекта. Простые расчеты показывают, что в беккроссном потомстве новые гомозиготы сильных по вредности генов не могут появиться, а гомозиготы субвитальных генов, если они не были элиминированны у выживших гомозиготных андрогенов, подавляются комплексом благоприятных генов, доставшихся от исходного гибрида. Вот поэтому гетерозис сохраняется во всех беккроссных поколениях (фиг. 3). С первыми и последующими беккроссными поколениями поступают точно так же, как и с исходным гибридом (фиг. 2). Дальнейшие беккроссы приводят, во-первых, к почти полному удалению деталей и полулеталей из генотипа гибрида и, во-вторых, к сохранению той численно преобладающей части генов, которые обеспечили гетерозис в исходном гибриде. После 5 или 6 беккроссов очищенный от вредных генов гибрид массово размножают путем внутригибридного скрещивания. У потомства, полученного в результате такого размножения, гетерозис не только сохраняется на уровне исходного гибрида, но и даже несколько усиливается (фиг. 4), что свидетельствует о полном решении проблемы закрепления гетерозиса у тутового шелкопряда. Полная общность генетических основ гетерозиса и его затухания у животных и растений позволяет данное изобретение рекомендовать для закрепления гетерозиса у сельскохозяйственных растений, у которых возможно получение от гибридов абсолютно гомозиготных особей андрогенного происхождения. Их получают путем стимулирования эмбрионального развития гаплоидной пыльцы с последующим превращением ее зародышевых клеток в диплоидные, развивающиеся в жизнеспособные фертильные растения. Методика варьирует в зависимости от биологических особенностей культуры. Графические материалы. Фиг. 1 А. Показана прямая зависимость между урожаем коконов тутового шелкопряда - основной показатель гетерозиса (1) и уровнями гетерозиготности (2) генетических вариантов гибрида неочищенных от леталей и полулеталей. Показатели урожая и гетерозиготности исходного гибрида первого варианта (1) приняты за 100%. Б. Показано полное отсутствие зависимости между урожаем коконов (1) и уровнями гетерозиготности (2) у генетических вариантов, очищенных от леталей и полулеталей. Это доказывает несостоятельность гипотезы гетерозиса "сверхдоминирования" и возможность сохранения гетерозиса в беккроссных поколениях. Фиг. 2. Схема очищения гибридов тутового шелкопряда от рецессивных леталей и полулеталей посредством возвратных скрещиваний гибридов с полученными от них абсолютно гомозиготными самцами А и Б породы. F 1 , F 2 - гибрид первого и второго поколения. F b1 , F b2 - первое и второе беккроссное поколение. Фиг. 3. Жизнеспособность исходного гибрида (1) и беккросных поколений (II), полученных по схеме, представленной на фиг. 2. Фиг. 4. Демонстрирует показатели частоты вредных генов в гетерозиготном состоянии (1), массы кокона (2), жизнеспособности (3) у исходного гибрида (I) и трансформируемого гибрида после четырех последовательных возвратных скрещиваний с гомозиготными самцами (II), а также у трех последовательных инбредных поколений (III-V). Выкормка каждого генетического варианта проводилась одновременно с контрольным партеногенетическим гибридом, показатели которого принимались за 100%. Во всех генетических вариантах гетерозис выше, чем у исходного гибрида, что свидетельствует о кардинальном решении проблемы закрепления гетерозиса. Стабильное сохранение гетерозиса во всех беккроссных поколениях уже свидетельствовало о принципиальной эффективности разрабатываемого способа. Но беккроссные поколения не применимы в практике из-за сложности их получения. Поэтому в заключительном эксперименте на тутовом шелкопряде изучали возможность закрепления гетерозиса уже не в беккроссных, а в нормальных поколениях. В этом заключительном опыте исходный гибрид вначале подвергался четырем беккроссам с гомозигонтыми самцами. В результате частота гетерозигот по леталям и полулеталям снизилась до 6,2% со 100% в исходном материале. Далее беккроссные поколения размножали инбридингом. Каждое инбредное поколение получали посредствам скрещивания брата с сестрой в пределах каждой отдельно взятой семьи. В результате частота вредных генов, погашенных нормальными аллелями, снизилась в первом инбредном поколении до 4,7, а во втором и третьем - до 3,5 и 2,6% соответственно. Инбредное размножение исключительно пагубно влияет на все хозяйственные показатели нормального инбредного потомства. Но в нашем опыте оно не только не оказало угнетающего действия на инбредное потомство, а, наоборот, привело к повышению у него средней массы одного кокона и жизнеспособности по сравнению с исходным, контрольным гибридом (фиг. 4). Следовательно, проблема закрепления гетерозиса у гибридов последующих поколений кардинально решена. БИБЛИОГРАФИЧЕСКИЕ ДАННЫЕ
1. Инге-Вечтомов С. И. 1989. Генетика с основами селекции. М. "Высшая школа", на стр. 557. 2. Хатт Ф. 1969. Генетика животных. Пер. с англ. под ред. д-ра биол. наук Я.Л. Глембоцкого. М., "Колос", на стр. 322. 3. Струнников В. А. 1998. Клонирование животных: теория и практика. - Природа, N 7, с.3 -9. 4. Струнников В.А. 1987. Генетические методы селекции и регуляции пола тутового шелкопряда. М. ВО "Агропромиздат", на стр. 35. 5. Струнников В.А. 1994. Природа гетерозиса и новые методы его повышения. - М. Наука, 108 с.
ФОРМУЛА ИЗОБРЕТЕНИЯ
1. Способ закрепления гетерозиса гибрида в последующих поколениях, включающий использование возвратных скрещиваний с абсолютно гомозиготными самцами, отличающийся тем, что для сохранения в генотипе гибрида благоприятных генов, определяющих гетерозис, и одновременно для удаления леталей и полулеталей применяют возвратные скрещивания гибридов с полученными от них методом односпермичного андрогенеза андрогенетическими абсолютно гомозиготными самцами и затем после нескольких беккроссов переключают беккроссные поколения на обычное массовое бисексуальное размножение посредством внутригибридных скрещиваний. 2. Способ по п.1, отличающийся тем, что абсолютно гомозиготных самцов тутового шелкопряда получают методом односпермичного андрогенеза, выполняемого путем облучения яиц в теле самки - лучами в дозе 80 кр., последующего спаривания с самцами исходных индивидуальных гибридов и прогрева облученных осемененных яиц в возрасте 60 - 80 мин в воде, нагретой до 38 o C, в течение 210 мин. 3. Способ по п.1, отличающийся тем, что, с целью резкого повышения гетерозиса гибридов промышленного назначения, закрепляют гетерозис только у возникших от двух родителей индивидуальных гибридов, проявивших максимальный гетерозис по сравнению с другими одновременно испытуемыми гибридами. 4. Способ по п.1, отличающийся тем, что способ закрепления гетерозиса применяют на гибридах сельскохозяйственных растений, у которых возможно получение андрогенетических абсолютно гомозиготных индивидуумов известными для каждого вида варьирующими методами стимулирования пыльцы к эмбриональному развитию и превращения развивающихся из нее зародышевых клеток в диплоидные абсолютно гомозиготные клетки, развивающиеся в фертильные растения.Способ предназначен для использования в сельском хозяйстве. Цель - значительное повышение эффективности гибридизации сельскохозяйственных культур. Гетерозис гибридов проявляется только в первом поколении. Установлено, что затухание гетерозиса в последующих поколениях гибрида в основном происходит из-за перехода рецессивных леталей, полулеталей и субвиталей в гомозиготное состояние и нарушения комплекса благоприятных скоординировано действующих генов. Устранение этих явлений приводит к закреплению гетерозиса в последующих поколениях. Оно выполняется посредством возвратных скрещиваний гибрида с искусственно полученными от него абсолютно гомозиготными андрогенетическими сыновьями, после чего генетически трансформируемый гибрид практически полностью очищается от вредных генов и одновременно сохраняет в целости комплекс благоприятных генов, определяющих гетерозис. Это позволяет полностью сохранить гетерозис в последующих уже промышленных поколениях, полученных в результате несложных внутригибридных скрещиваний, что доказано экспериментами на тутовом шелкопряде. Способ также предназначен для сельскохозяйственных растений, у которых возможно получение андрогенетических абсолютно гомозиготных особей. 3 з.п.ф-лы, 4 ил.
Изобретение относится к способам, используемым в сельском хозяйстве. Общеизвестен естественный способ сохранения гетерозиса в ряду последующих поколений посредством вегетативного размножения у растений, дополнительно обладающих половым размножением. Многочисленные исследования в этой области на других растениях, не способных к вегетативному размножению, и животных не увенчались полным успехом (1), потому что природа гетерозиса все еще оставалась большой загадкой генетики (2). В литературе не было высказано даже сколько-нибудь реальных теоретических подходов к кардинальному решению этой важной проблемы. У некоторых животных гетерозис можно закрепить клонированием. Однако этим способом пока получают единицы идентичных матери потомков. У тутового шелкопряда клонирование разработано более успешно, но для практического использования в плане сохранения гетерозиса оно не приемлемо по двум причинам: из-за большой трудоемкости массового получения партеногенетического потомства и меньшей, по сравнению с самцами, продуктивности женского пола, из которого состоят клоны (3). Перспективные результаты получены авторами после того, как они разработали метод мейотического партеногенеза и получили от партеногенетических клонов абсолютно гомозиготных самцов тутового шелкопряда (4). Их возвратное скрещивание с партеногенетическим клоном гибридного происхождения позволило закреплять гетерозис в беккроссных поколениях (5). Но это было открытием лишь принципиальной возможности закрепления гетерозиса. Практического же значения этот метод не имел и, следовательно, не мог быть запатентован в качестве способа. Это объяснялось тем, что гомозиготных самцов могли получать только от высокожизнеспособных женских партеноклонов с высокой склонностью к партеногенезу. У коммерческих пород и гибридов абсолютных гомозиготов практически получать не удавалось, поэтому мейотический партеногенез был использован только для рекогносцировочных опытов, направленных на выяснение возможности решения проблемы. Изобретение способа закрепления гетерозиса у тутового шелкопряда, пригодного для производства, стало возможным после открытия авторами односпермичного андрогенеза (1998, неопубликовано). Сущность изобретения. Гетерозис проявляется только в первом поколении гибрида. В последующих поколениях, начиная со второго, он резко затухает. Поэтому, чтобы вырастить гетерозисный гибрид приходится каждый раз вновь повторять межсортовую или межпородную гибридизацию. Этот процесс технически сложный и весьма трудоемкий, а применительно ко многим растительным культурам он просто невыполним, хотя их гибриды, если бы они были получены, дали бы удивительно высокие урожаи по сравнению с родительскими формами. Примером этого служат многие сельскохозяйственные растения. Эти проблемы были бы кардинально решены, если бы удалось разработать эффективный способ закрепления гетерозиса в последующих поколениях. Такой способ одновременно открыл бы совершенно новый подход к созданию еще более выдающихся по гетерозису гибридов. Известно, что любой промышленный гибрид получают посредством скрещивания огромной массы индивидуумов двух родительских форм. А эти индивидуумы весьма дифференцированы по комбинационной способности. Поэтому производство довольствуется средним гетерозисом по всем вместе взятым индивидуальным гибридам, каждый из которых происходит от половых клеток двух родителей. В то время как редкие индивидуальные гибриды обладают поистине фантастическим гетерозисом, в следующем поколении он безвозвратно теряется. Предлагаемый способ позволит закреплять этот мощный гетерозис в последующих поколениях гибрида и размножать его в неограниченных количествах. Одной из причин гетерозиса считали благоприятное действие на развитие и жизнедеятельность организма гетерозиготности вообще всех генов, независимо от их специфичности (гипотеза "сверхдоминирования"). Авторы на тутовом шелкопряде экспериментально доказали, что гетерозис возникает в результате двух главных причин. Первая - интеграция в генотипе гибридов большого количества скоординированных в своем действии благоприятных генов, контролирующих жизнеспособность. Вторая - переход в гетерозиготное состояние не всех генов генотипа, а только рецессивных деталей, полулеталей и субвиталей (4). На фиг. 1 приведены доказательства этого. Следовательно, снижение гетерозиса в последующих поколениях гибридов в основном объясняется неизбежным при скрещивании гибрида в его пределах переходом части рецессивных деталей и полулеталей в гомозиготное состояние и нарушением в процессе мейоза комплекса благоприятных генов, повышающих жизнеспособность. Поэтому авторы пришли к выводу, что закрепить гетерозис в последующих поколениях можно, если в генотипе гибрида полностью сохранить или даже улучшить комплекс всех благоприятных генов и практически полностью удалить из генотипа рецессивные летали и полулетали. Эта задача решена авторами следующим способом. В качестве исходного материала выбирают генетически отдаленные две породы, от скрещивания которых возникают наиболее высокогетерозисные гибриды. От этих двух пород получают серию индивидуальных гибридов, каждый из которых происходит только от двух родителей. Путем сравнительных испытаний выбирают 10 лучших по гетерозису индивидуальных гибридов. От каждого гибрида получают абсолютно гомозиготных потомков методом односпермического андрогенеза, выполнение которого доступно селекционерам. Для этого неосемененных самок любой породы облучают - лучами в дозе 80 кр. Затем самки спариваются с самцами индивидуальных гибридов. Отложенные яйца в возрасте 60-80 минут после откладки при температуре 25 o C прогревают 210 минут в воде, нагретой до 38 o C. Абсолютные гомозиготы в подавляющем большинстве погибают на разных стадиях развития из-за того, что в гаплоидном генотипе, доставшимся им от отца, содержится много летальных, полулетальных и субвитальных генов. При диплоидизации ядра пронуклеуса они переходят в гомозиготное состояние, чаще всего несовместимое с нормальным развитием организма. Выживают только те гомозиготы, которым в ходе мейоза не досталось или досталось, но очень мало, вредных генов, в основном слабого действия (5). Выращенных абсолютно гомозиготных особей возвратно (беккросс) скрещивают с исходным гибридом, получая, таким образом, первое беккроссное поколение (фиг. 2). Созревание исходного гибрида и абсолютных гомозиготов должно быть синхронизировано путем задержки начала выращивания первого на время, равное продолжительности цикла развития взятого объекта. Простые расчеты показывают, что в беккроссном потомстве новые гомозиготы сильных по вредности генов не могут появиться, а гомозиготы субвитальных генов, если они не были элиминированны у выживших гомозиготных андрогенов, подавляются комплексом благоприятных генов, доставшихся от исходного гибрида. Вот поэтому гетерозис сохраняется во всех беккроссных поколениях (фиг. 3). С первыми и последующими беккроссными поколениями поступают точно так же, как и с исходным гибридом (фиг. 2). Дальнейшие беккроссы приводят, во-первых, к почти полному удалению деталей и полулеталей из генотипа гибрида и, во-вторых, к сохранению той численно преобладающей части генов, которые обеспечили гетерозис в исходном гибриде. После 5 или 6 беккроссов очищенный от вредных генов гибрид массово размножают путем внутригибридного скрещивания. У потомства, полученного в результате такого размножения, гетерозис не только сохраняется на уровне исходного гибрида, но и даже несколько усиливается (фиг. 4), что свидетельствует о полном решении проблемы закрепления гетерозиса у тутового шелкопряда. Полная общность генетических основ гетерозиса и его затухания у животных и растений позволяет данное изобретение рекомендовать для закрепления гетерозиса у сельскохозяйственных растений, у которых возможно получение от гибридов абсолютно гомозиготных особей андрогенного происхождения. Их получают путем стимулирования эмбрионального развития гаплоидной пыльцы с последующим превращением ее зародышевых клеток в диплоидные, развивающиеся в жизнеспособные фертильные растения. Методика варьирует в зависимости от биологических особенностей культуры. Графические материалы. Фиг. 1 А. Показана прямая зависимость между урожаем коконов тутового шелкопряда - основной показатель гетерозиса (1) и уровнями гетерозиготности (2) генетических вариантов гибрида неочищенных от леталей и полулеталей. Показатели урожая и гетерозиготности исходного гибрида первого варианта (1) приняты за 100%. Б. Показано полное отсутствие зависимости между урожаем коконов (1) и уровнями гетерозиготности (2) у генетических вариантов, очищенных от леталей и полулеталей. Это доказывает несостоятельность гипотезы гетерозиса "сверхдоминирования" и возможность сохранения гетерозиса в беккроссных поколениях. Фиг. 2. Схема очищения гибридов тутового шелкопряда от рецессивных леталей и полулеталей посредством возвратных скрещиваний гибридов с полученными от них абсолютно гомозиготными самцами А и Б породы. F 1 , F 2 - гибрид первого и второго поколения. F b1 , F b2 - первое и второе беккроссное поколение. Фиг. 3. Жизнеспособность исходного гибрида (1) и беккросных поколений (II), полученных по схеме, представленной на фиг. 2. Фиг. 4. Демонстрирует показатели частоты вредных генов в гетерозиготном состоянии (1), массы кокона (2), жизнеспособности (3) у исходного гибрида (I) и трансформируемого гибрида после четырех последовательных возвратных скрещиваний с гомозиготными самцами (II), а также у трех последовательных инбредных поколений (III-V). Выкормка каждого генетического варианта проводилась одновременно с контрольным партеногенетическим гибридом, показатели которого принимались за 100%. Во всех генетических вариантах гетерозис выше, чем у исходного гибрида, что свидетельствует о кардинальном решении проблемы закрепления гетерозиса. Стабильное сохранение гетерозиса во всех беккроссных поколениях уже свидетельствовало о принципиальной эффективности разрабатываемого способа. Но беккроссные поколения не применимы в практике из-за сложности их получения. Поэтому в заключительном эксперименте на тутовом шелкопряде изучали возможность закрепления гетерозиса уже не в беккроссных, а в нормальных поколениях. В этом заключительном опыте исходный гибрид вначале подвергался четырем беккроссам с гомозигонтыми самцами. В результате частота гетерозигот по леталям и полулеталям снизилась до 6,2% со 100% в исходном материале. Далее беккроссные поколения размножали инбридингом. Каждое инбредное поколение получали посредствам скрещивания брата с сестрой в пределах каждой отдельно взятой семьи. В результате частота вредных генов, погашенных нормальными аллелями, снизилась в первом инбредном поколении до 4,7, а во втором и третьем - до 3,5 и 2,6% соответственно. Инбредное размножение исключительно пагубно влияет на все хозяйственные показатели нормального инбредного потомства. Но в нашем опыте оно не только не оказало угнетающего действия на инбредное потомство, а, наоборот, привело к повышению у него средней массы одного кокона и жизнеспособности по сравнению с исходным, контрольным гибридом (фиг. 4). Следовательно, проблема закрепления гетерозиса у гибридов последующих поколений кардинально решена. БИБЛИОГРАФИЧЕСКИЕ ДАННЫЕ 1. Инге-Вечтомов С. И. 1989. Генетика с основами селекции. М. "Высшая школа", на стр. 557. 2. Хатт Ф. 1969. Генетика животных. Пер. с англ. под ред. д-ра биол. наук Я.Л. Глембоцкого. М., "Колос", на стр. 322. 3. Струнников В. А. 1998. Клонирование животных: теория и практика. - Природа, N 7, с.3 -9. 4. Струнников В.А. 1987. Генетические методы селекции и регуляции пола тутового шелкопряда. М. ВО "Агропромиздат", на стр. 35. 5. Струнников В.А. 1994. Природа гетерозиса и новые методы его повышения. - М. Наука, 108 с.
Формула изобретения
1. Способ закрепления гетерозиса гибрида в последующих поколениях, включающий использование возвратных скрещиваний с абсолютно гомозиготными самцами, отличающийся тем, что для сохранения в генотипе гибрида благоприятных генов, определяющих гетерозис, и одновременно для удаления леталей и полулеталей применяют возвратные скрещивания гибридов с полученными от них методом односпермичного андрогенеза андрогенетическими абсолютно гомозиготными самцами и затем после нескольких беккроссов переключают беккроссные поколения на обычное массовое бисексуальное размножение посредством внутригибридных скрещиваний. 2. Способ по п.1, отличающийся тем, что абсолютно гомозиготных самцов тутового шелкопряда получают методом односпермичного андрогенеза, выполняемого путем облучения яиц в теле самки - лучами в дозе 80 кр., последующего спаривания с самцами исходных индивидуальных гибридов и прогрева облученных осемененных яиц в возрасте 60 - 80 мин в воде, нагретой до 38 o C, в течение 210 мин. 3. Способ по п.1, отличающийся тем, что, с целью резкого повышения гетерозиса гибридов промышленного назначения, закрепляют гетерозис только у возникших от двух родителей индивидуальных гибридов, проявивших максимальный гетерозис по сравнению с другими одновременно испытуемыми гибридами. 4. Способ по п.1, отличающийся тем, что способ закрепления гетерозиса применяют на гибридах сельскохозяйственных растений, у которых возможно получение андрогенетических абсолютно гомозиготных индивидуумов известными для каждого вида варьирующими методами стимулирования пыльцы к эмбриональному развитию и превращения развивающихся из нее зародышевых клеток в диплоидные абсолютно гомозиготные клетки, развивающиеся в фертильные растения.
Подробное решение параграф § 32 по биологии для учащихся 10 класса, авторов Сивоглазов В.И., Агафонова И.Б., Захарова Е.Т. 2014
Вспомните!
Что такое селекция?
Приведите примеры известных вам пород животных и сортов растений.
Сорта яблок Антоновка, груша Северянка, породы собак: ротвейлер, карликовый пудель, колли.
Вопросы для повторения и задания
1. Что такое селекция?
Селекция (от лат. selectio - отбор) - наука о создании новых и улучшении существующих сортов растений, пород животных и штаммов микроорганизмов. Одновременно под селекцией понимают и сам процесс создания сортов, пород и штаммов. Теоретической основой селекции является генетика.
2. Что называют породой, сортом, штаммом?
Порода, сорт или штамм - это совокупность особей одного вида, искусственно созданная человеком и характеризующаяся определёнными наследственными свойствами.

5. Какие сложности возникают при постановке межвидовых скрещиваний?
Отдалённая гибридизация заключается в скрещивании разных видов. В растениеводстве с помощью отдалённой гибридизации создана новая зерновая культура - тритикале, гибрид ржи с пшеницей. Эта культура сочетает многие свойства пшеницы (высокие хлебопекарные качества) и ржи (способность расти на бедных песчаных почвах). Классическим примером межвидовых гибридов в животноводстве является мул, полученный при скрещивании осла с кобылицей, который значительно превосходит родителей по выносливости и работоспособности. В Казахстане при скрещивании диких горных баранов-архаров с тонкорунными овцами была создана знаменитая архаромериносная порода овец. Однако применение межвидовых скрещиваний имеет определённые сложности, потому что получаемые гибриды часто оказываются бесплодными (стерильными) или низкоплодовитыми. Стерильность гибридов связана с отсутствием у них парных гомологичных хромосом. Это делает невозможным процесс конъюгации. Следовательно, мейоз не может завершиться, и половые клетки не образуются.
6. Получают ли и используют ли в вашем регионе межвидовые гибриды? Используя дополнительные источники информации, выясните, гибридами каких видов являются такие организмы, как бестер, хонорик, лошак, рафанобрассика. Какой интерес представляют они для сельского хозяйства?

Подумайте! Вспомните!

2. Почему для каждого региона нужны свои сорта растений и породы животных? Какие сорта и породы характерны для вашего региона? В чём их особенности и преимущества?
Так как условия среды в различных регионах разные, и сорта и породы должны быть приспособлены к конкретным условиям. Особенности растениеводства Южного Урала

3. Из большого разнообразия видов животных, обитающих на Земле, человек отобрал для одомашнивания сравнительно немного видов. Как вы считаете, чем это объясняется?
Процесс одомашнивания диких животных начинается с искусственной селекции отдельных индивидов для получения потомства с определенными признаками, необходимыми человеку. Индивиды, как правило, выбираются в соответствии с определёнными желаемыми характеристиками, включая снижение агрессивности по отношению к человеку и представителям собственного вида. В этом отношении принято говорить об укрощении дикого вида. Целью одомашнивания является использование животного в сельском хозяйстве в качестве сельскохозяйственного животного или в качестве домашнего питомца. Если эта цель достигнута, можно говорить об одомашненном животном. Одомашнивание животного коренным образом изменяет условия для дальнейшего развития вида. Естественное эволюционное развитие заменяется искусственной селекцией по критериям разведения. Таким образом, в рамках одомашнивания меняются генетические свойства вида.
4. Гетерозис в последующих поколениях обычно не сохраняется, затухает. Почему это происходит?
При скрещивании разных пород животных или сортов растений, а также при межвидовых скрещиваниях в первом поколении у гибридов повышается жизнеспособность и наблюдается мощное развитие. Явление превосходства гибридов по своим свойствам родительских форм получило название гетерозиса, или гибридной силы. Проявляется в первом поколении, а во втором затухает.
5. Как вы думаете, почему лигры рождаются только в зоопарках и не встречаются в дикой природе? Объясните свою точку зрения.
Лигры - межвидовые гибриды между львом и тигрицей - выглядят как огромные львы с размытыми полосами. Следовательно, его родители относятся к одному и тому же биологическому роду пантер, но разным видам. Внешне он заметно отличается от своего противоположного гибрида, тигрольва. Является крупнейшим представителем семейства кошачьих, существующих в настоящее время. Выглядит как гигантский лев с размытыми полосами. Лигры не встречаются в природе главным образом потому, что в естественной среде львы и тигры почти не имеют шансов встретиться: современный ареал льва включает в основном центральную и южную Африку (хотя в Индии существует последняя уцелевшая популяция азиатских львов), в то время как тигр - исключительно азиатский вид. Поэтому скрещивание видов происходит, когда животные долгое время живут в одном вольере или клетке (например, в зоопарке или цирке), но потомство дают лишь 1-2 % пар, из-за чего в мире сегодня числится не более двух десятков лигров.
6. Как вы считаете, может ли применяться массовый отбор при разведении животных? Докажите свое мнение.
Не применяют. массовый отбор - это отбор по фенотипу. Индивидуальный - по генотипу. Производители у животных - особи с хорошо прописанной родословной т.е. генотип по нужным признакам достаточно хорошо известен. Да и особенности животных - необходимо время для достижения половой зрелости, небольшое число потомства (по сравнению с растениями - сейчас можно считать решенной проблемой - искусственное осеменение, суррогатные самки) и невозможность бесполого размножения.
7. Используя дополнительную литературу и ресурсы Интернета, подготовьте сообщение или презентацию об истории селекции с древних времён до настоящего времени.
Селекция как способ выведения пород домашних животных и сортов культурных растений существует издавна. Около 8000-9000 лет назад с появлением сельского хозяйства на Ближнем Востоке, а позже в Европе и Азии началось развитие растениеводства и животноводства. Уже с тех времен люди стали заниматься искусственным отбором с целью выведения пород животных и сортов растений с хозяйственно-ценными качествами. О первых селекционных мероприятиях, известных еще почти 6000 лет назад в Эламе (Двуречье), можно судить по изображе¬нию родословной лошадей, обнаруженной на печатке. Существуют также сведения, что арабы задолго до новой эры применяли искусственное опыление финиковых пальм. В Римской империи сохранились документы с подробным описанием приемов, используемых при разведении животных. В трудах ученых Древнего Китая и Древнего Рима имеются указания на значение отбора колосьев у злаков и даются рекомендации по проведению такого отбора.
На первых порах селекционные мероприятия ограничивались отбором. Он носил бессознательный характер, велся длительное время (10-15 лет). Селекционеры, не имея теоретической базы, руководствовались опытом и интуицией. Они учитывали полезные свойства родительских особей, но целенаправленно проводить селекцию не могли. Результаты скрещивания часто оказывались неожиданными, и в потомстве не обнаруживалось ожидаемого признака. Тем не менее, безвестные селекционеры оставили в наследство немало ценных сортов культурных растений и пород домашних животных. Например, ряд лучших сортов хлопчатника, возделываемых ныне в России и США, позаимствован у крестьян старых мексиканских деревень. Методом бессознательного отбора выведены сорта льна-долгунца в некоторых районах Пско¬ва: низкорослые растения шли на хозяйственные нужды, а семена высоких использовались на посев. Известны сорта озимой (например, Крымка, Полтавка, Сандомирка) и яровой (Улька, Гирка, Сыр-Бидай и др.) пшеницы с ценными хозяйственными качествами, выведенные в давние времена.
Однако отбор по хозяйственно-полезным признакам и свойствам без учета механизмов их наследуемости и изменчивости нередко давал нежелательные результаты. К примеру, отбор по экстерьеру тонкорунных овец на комолость приводил к появлению крипторхизма; избавление от пегости на шее у романовских овец ослабляло их жизнеспособность; повышение оброслости шерстью у овец сопровождалось снижением их веса. Не удавалось вывести и чистую линию виандоттов (порода кур) с розовидным гребнем; несмотря на выбраковывание цыплят с листовидным гребнем, они появлялись в потомстве. Очевидно, порода состояла из генерозигот по этому гену, так как гомозиготы обладали сниженной плодовитостью.
Все это свидетельствовало о том, что желаемый ре¬зультат нельзя получить без теоретических знаний. С конца XVIII - начала XIX в. работы селекционеров носили уже научный характер. Главной задачей селекции стало изучение генетики таких признаков, как продуктивность животных и урожайность растений. Разрешение задач селекции невозможно без знаний, касающихся генетического анализа, т. е. без знаний типа наследования признаков (доминантный или рецессивный), типа доминирования, характера наследования (аутосомное или сцепленное с полом, независимое или сцепленное), типа и характера взаимодействия генов в онтогенезе. Главное внимание селекционеры должны уделять проблемам взаимоотношения генотипа и среды, ибо от факторов последней во многом зависит экспрес¬сивность и пенетрантность изучаемых признаков.
8. Существуют ли в вашем регионе селекционные станции или центры? Какие исследования они проводят? Каковы их достижения? Вместе с учителем организуйте экскурсию на такую станцию.
Ю-У НИИ плодоовощеводства и картофелеводства, г. Челябинск
Эх, яблочко, ранеточка...
В 1931 году по инициативе И. В. Мичурина было создано первое на Южном Урале научно-исследовательское учреждение по садоводству - Уральская зональная плодово-ягодная опытная станция. Организатором этой станции был Валерий Павлович Ярушин.
А уже в следующем году начались научные исследования по селекции и подбору сортов, пригодных для выращивания в суровых условиях Челябинской и Курганской областей, в том числе и входивших в них тогда Камышловском и Каменск-Уральском районах ныне Свердловской области. Ученые приступили к обследованию и сбору лучших форм плодово-ягодных культур восточнее Уральского хребта. В 1934 году сотрудники станции зарегистрировали крупный массив дикорастущей вишни - 2270 га - в Карагайской лесной даче (Анненский бор). В том же году научная экспедиция, возглавляемая доктором сельскохозяйственных наук Э. П. Сюбаровой (БелНИИ плодоводства) и челябинским ученым М. Н. Саламатовым, обследовала заросли дикой степной вишни в Верхнеуфалейском и Полтавском районах Челябинской области. Одновременно были завезены обширные коллекции сливы с Дальнего Востока, из Канады, Северной Америки, средней полосы России, Поволжья, карзинские сливы из Сибири. Из отобранного материала в 1937 году ученые выделили сорта и предложили первый уральский сортимент ягодных культур.
В те годы, как в народе, так и агрономической науке, считалось, что из-за сурового климата с морозными и продолжительными зимами, садоводство на Урале невозможно. Южноуральским ученым потребовалось всего двадцать лет - по селекционным меркам весьма небольшой срок, чтобы опровергнуть это широко распространенное мнение. Благодаря селекции и изучению новых сортов плодово-ягодных культур на челябинской опытной станции, у нас стало стремительно развиваться садоводство.
Первые коллективные сады в области появились вскоре после войны. В 1948 году образовались "Тракторосад", "Дружба" в Металлургическом районе, "Локомотив" в Советском районе, сады Магнитогорского металлургического комбината. Их появлению предшествовала длительная и вязкая борьба в коридорах тогдашней власти с противниками создания таких садов. Тем не менее, коллективное садоводство развивалось и развивается до сих пор. В настоящее время садоводы производят основную часть плодоовощной продукции.
К началу 50-х годов нашими учеными были собраны и изучались 442 сортообразца яблони, в том числе 210 сортов урало-сибирской селекции. Первые официально оформленные авторскими свидетельствами сорта плодово-ягодной опытной станции были именно яблони. Основой довоенного сортимента стали местные ранетки. Они отличаются приспособленностью к уральскому и сибирскому климату, высокой урожайностью и малыми размерами плодов весом от 15 до 50 г. Старшее поколение еще помнит эти яркие красивые яблочки-ранетки - Любимец, Анисик омский, Пониклое. Они казались тогда пределом мечтаний, особенно для детворы. Но к 60-м годам в активе института было уже 25 районированных сортов:14 яблонь, 4 груши, 4 сливы и 3 ягодных культуры.
В 1964 году Уральская зональная плодово-ягодная опытная станция была переименована в Челябинскую плодоовощную селекционную станцию им. И. В. Мичурина. К тому времени в области уже успешно работал специализированный трест "Плодопром", который возглавлял Всеволод Иванович Назаров. Тесное сотрудничество науки и производства, увлеченность и энтузиазм их сотрудников подняли южноуральское садоводство на невиданные высоты. При поддержке руководителей области были созданы плодовые питомники: "Смолинский" в пригороде Челябинска, "Мичуринский" в Карталах, "Радужный" в Магнитогорске, "Тюбелясский" в горнозаводской зоне. При Смолинском и Мичуринском плодопитомниках были организованы государственные сортоиспытательные участки.
Именно тогда, в 50-60-е годы в колхозах и совхозах, находящихся при МТС, закладывались небольшие сады от 20 до 100 га. Некоторые из них сохранились до сих пор. На пике развития садоводства области общественные сады занимали у нас площадь 8 тысяч гектаров, а сама отрасль была в целом прибыльной.
В пятидесятые годы в нашей области началась также работа по научному обеспечению картофелеводства. К началу 80-х годов Челябинская плодоовощная селекционная станция им. И. В. Мичурина имела в своем активе 29 собственных сортов плодово-ягодных культур и картофеля, занесенных в Государственный реестр селекционных достижений, допущенных к использованию.
Ученые станции начали использовать в селекции растений мировую коллекцию форм плодово-ягодных культур и картофеля из Всесоюзного института растениеводства и других научно-исследовательских учреждений, в том числе зарубежных. В селекционный процесс вовлекался генетический фонд, внедрялись новые селекционные технологии, позволяющие сокращать селекционный процесс, возросла подготовка кадров высшей квалификации. Научные разработки южноуральских ученых стали внедряться в хозяйствах Челябинской, Курганской, Кустанайской, Оренбургской и других областях, в Республике Башкортостан.
Новое имя - новые цели
В ноябре 1991-го по решению правительства РФ Челябинская плодоовощная опытная станция им. И. В. Мичурина была преобразована в Южно-Уральский НИИ плодоовощеводства и картофелеводства. Изменилось не только название. Перед институтом были поставлены более серьезные цели и направления научно-исследовательской деятельности. Помимо селекции плодово-ягодных культур и картофеля, жизнь потребовала создания новых ресурсосберегающих экологически чистых технологий селекции и возделывания этих культур, научных исследований по гибридизации, а также производства элитных саженцев новых перспективных сортов садовых культур и семян картофеля на оздоровленной, безвирусной основе с применением биотехнологии.
За весь период деятельности института, начиная с Уральской зональной плодово-ягодной станции, челябинскими селекционерами создано более 200 сортов плодово-ягодных культур, 18 сортов картофеля, разработаны технологии промышленного и любительского садоводства, технологии производства картофеля с урожайностью до 70 т/га.
В государственный реестр селекционных достижений, допущенных к использованию, в разные годы было внесено 110 сортов. Об уровне научно-исследовательской работы говорит тот факт, что на сегодняшний день институт имеет более 100 авторских свидетельств и патентов на сорта и изобретения.
Впервые на Южном Урале созданы модели интенсивного сорта плодовых, ягодных культур, картофеля до 2020 года, разработаны схемы селекции на продуктивность, зимостойкость, в т.ч. устойчивость цветков к весенним заморозкам, качество продукции, иммунитет. Институт располагает богатым генетическим фондом садовых культур, который насчитывает 64 тысячи гибридных растений, в т.ч. 39 - плодовых, 25 тысяч сеянцев ягодных культур. Объемы гибридных скрещиваний составляют 45 тысяч цветков.
Научные исследования по селекции и агротехнике садовых культур и картофеля ведутся в творческом содружестве с ведущими научно-исследовательскими институтами и опытными станциями России, ближнего и дальнего зарубежья.
На сегодняшний день институт имеет селекционный сад площадью более 100 гектаров. Именно здесь сосредоточена основная научная деятельность южноуральских селекционеров.
Яблоня. Впервые в мировой практике садоводства селекционеры института по оригинально разработанной методике вывели сорта естественных карликов с высотой деревьев в 1,5-2,5 м. При размножении их на клоновых вегетативно размножаемых карликовых подвоях, они становятся естественными стланцами (0,8-1,5 м).
Груша - удивительная культура на Урале. Ежегодно плодоносит. Плоды вкусные, сладкие, пригодные для переработки. Привлечение в селекцию отборных форм уссурийской груши позволило создать сорта, отличающиеся высокой зимостойкостью и продуктивностью, высокими вкусовыми качествами плодов, моногенной устойчивостью к парше и полевой устойчивостью к грушевому галловому клещу.
Абрикос и слива. Выведены местные сорта и выделены отборные формы, отличающиеся повышенной зимостойкостью плодовых почек и высоким качеством плодов.
Вишня. Накоплены экспериментальные данные для создания генотипов, устойчивых к коккомикозу. Продолжается выделение доноров с геном моноустойчивости к коккомикозу и выявление форм степной вишни с полевой устойчивостью к коккомикозу. С этой целью и для пополнения коллекции проведены экспедиции по обследованию дикоросов вишни на территории Башкирии, Челябинской и Курганской областей. Отобраны формы степной и лесной вишни, устойчивые к коккомикозу, крупноплодные, хорошим вкусом плодов.
Ягодные культуры. Ведутся исследования по селекции новых сортов, устойчивых к неблагоприятным факторам среды, высокозимостойких, с повышенной устойчивостью цветков к весенним заморозкам, высокопродуктивных, с отличным качеством плодов. Составлены модели оптимального сорта ягодных культур с учетом технологических запросов селекции 2020-2025 годов. Разработаны технологии возделывания шиповника, крыжовника, жимолости, смородины, технология размножения смородины в пленочных теплицах.
Вернуть былую славу
Реформы, проводимые в аграрном секторе России с начала 90-х годов, оказались разрушительными для садоводства. Как отрасль народного хозяйства оно фактически перестало существовать. Только в Челябинской области доля промышленного садоводства с 65% снизилась до 1%. Почти не осталось плодоносящих насаждений, опытно-производственные плодопитомники практически прекратили свою деятельность, а их продукция удовлетворяет спрос рынка всего на 12-15%.
В области отсутствует целевая программа развития промышленного садоводства, основанная на достоверных материалах инвентаризации насаждений и новых разработках научных учреждений последних лет.
В этом нет вины науки. Ученые Южно-Уральского научно-исследовательского института плодоовощеводства и картофелеводства не раз предлагали сформировать научные основы возрождения отрасли. Однако состояние макроэкономики и чиновничий консерватизм не дают возможности продвинуться вперед. Расчет на то, что коллективное любительское садоводство (т.е. частный сектор) восполнит потери промышленного садоводства, не оправдывается. Да и не может оправдаться. Не смотря на то, что ЮУНИИПОК ежегодно производит более 60 тысяч саженцев плодовых и ягодных культур для населения, частный сектор страдает от недостатка посадочного материала, особенно новых сортов, потребность в котором удовлетворяется на 47-50%. Эту нишу пытаются заполнить мелкие производители саженцев - частники. Но качество их посадочного материала часто не выдерживает никакой критики по всем параметрам.
Реально ли возродить общественное садоводство в новых экономических условиях?
Вполне, считают южноуральские ученые и селекционеры. Но для этого на государственном и областном уровне, по их мнению, необходимо решить самые насущные вопросы:
С площадей под садами от начала подготовки территорий и до вступления в плодоношение насаждений отменить налог на землю
На возвратной основе выделять капитальные вложения для посадки многолетних насаждений
На промышленных предприятиях области организовать производство специальной техники для садоводства
Необходимы законы по защите отечественного производителя на собственном рынке
Стимулировать малый бизнес для организации переработки продукции садоводства
В системе профтехобразования необходимо организовать подготовку кадров для садоводства
Весьма актуально и объединение научного потенциала уральских ученых-аграрников. Для этих целей необходимо создание Уральского научно-методического центра с размещением его в Челябинске. Польза от такого объединения огромная. Доказательством тому координационный совет по картофелю, созданный в апреле 2000 года по инициативе ГНУ ЮУНИИПОК на общественных началах. За 8 лет совместной работы институты картофелеводства провели мощную мобилизацию генофонда, пополнили коллекции, договорились о комбинациях скрещивания, регулярно обмениваются информацией, исходным и селекционным материалом. Практика координационного совета показала, что эта форма работы чрезвычайно эффективна, жизнеспособна и поэтому заслуживает развития и совершенствования.
Начата работа и по объединению садоводческих учреждений. Подписано многостороннее соглашение между шестью научными учреждениями Уральского и прилегающих к нему регионов - ГНУ ЮУНИИПОК, БашНИИСХ, Удмуртский НИИСХ, Костанайский НИИСХ, Казахский НИИКОХ, Карабалыкская опытная станция.
Аграрная наука сегодня переживает трудные времена, но она жива и готова внести свой вклад в возрождение сельскохозяйственного производства, в том числе и южноуральского промышленного садоводства.



